Noyau
Ce que vous savez du lycée
| De la seconde* : | / |
| De la première** : | Les chromosomes sont des structures universelles aux cellules eucaryotes (organismes dont les cellules ont un noyau). Chaque chromosome est constitué d’une longue molécule d’ADN associée à des protéines structurantes. |
| De la terminale*** : | / |
**adapté du programme officiel de Sciences de la Vie et de la Terre de première générale publié au Bulletin Officiel de l’Education Nationale.
***adapté du programme officiel de spécialité de Sciences de la Vie et de la Terre de terminale générale publié au Bulletin Officiel de l’Education Nationale.
1. Présentation
1.1. Définition
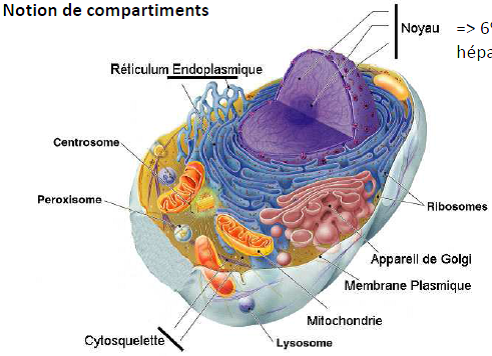
- Compartiment subcellulaire protubérant contenant de l’ADN organisé en chromosome.
=> 6% du volume d’une cellule hépatique.
- Site principal de la synthèse d’ADN et d’ARN.
Quel est l’intérêt de compartimentaliser ?
=> spécialisation compartimentale pour mettre en contact spécifique des partenaires moléculaires
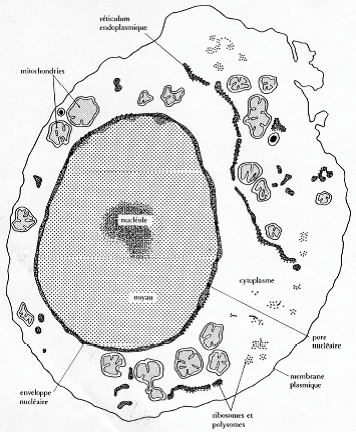
1.2. Mise en évidence
Photographie au microscope à fluorescence, coloration au DAPI (intercalant de l’ADN).
Photographie au MET.
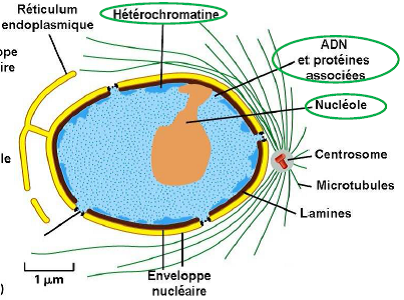
Dessin représentatif d’une cellule d’un embryon de souris.
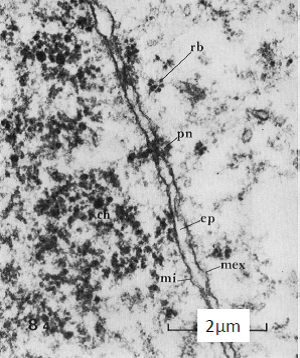
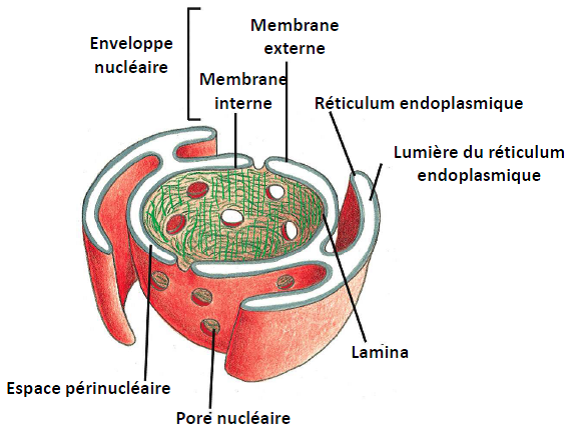
1.3. Enveloppe Nucléaire
Double membrane (membrane interne et externe) entourant le noyau et perforée de pores nucléaires.
Où sont le cytoplasme et le nucléoplasme ?
=> le cytoplasme est du côté de la membrane externe et le noyau est du côté de la membrane interne
Photographie au MET, cellule de mammifère, vue transversale.
1.4. Pores Nucléaires
Canal aqueux au travers de l’enveloppe nucléaire pour le passage de certaines molécules du noyau au cytoplasme et inversement.
Complexe du pore nucléaire :
Structure multi-protéique qui forme le pore
=> composé de protéines : les nucléoporines.
Mammifères : ~ 3500 pores par enveloppe nucléaire.
Photographie au MET après cryofracture, cellule de mammifère.
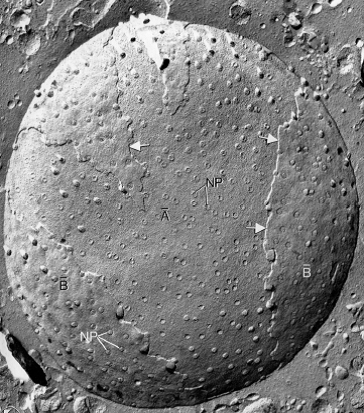
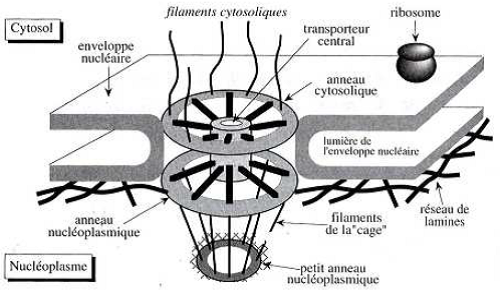
Schéma d’une vue latérale
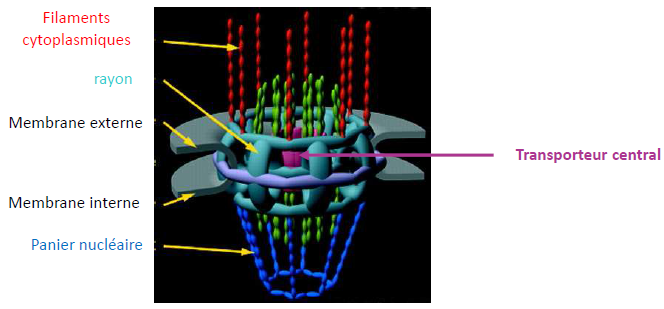
Structure du pore nucléaire
- 8 rayons qui se projettent radialement depuis la paroi du pore et entourant le transporteur central.
- 8 filaments cytoplasmiques attachés à l’anneau cytoplasmique.
- Des filaments nucléaires attachés à l’anneau nucléoplasmique et se rejoignant pour former le panier.

CF: filaments cytoplasmiques
NB: Nuclear Basket (panier nucléaire)
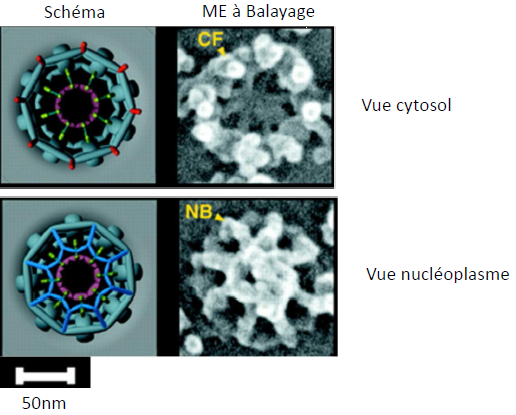
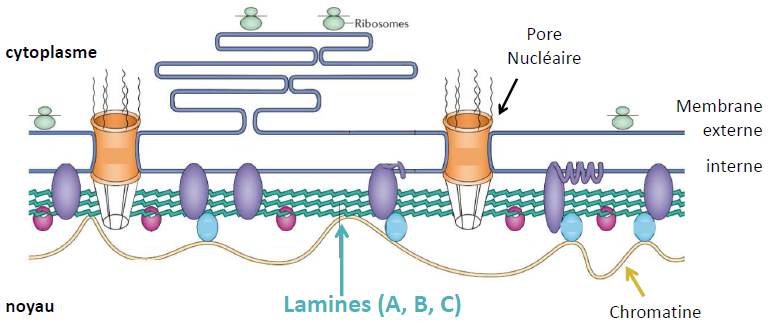
1.5. Lamina
= Filament intermédiaire (cf cours cytosquelette) qui tapisse la surface interne de l’enveloppe nucléaire.
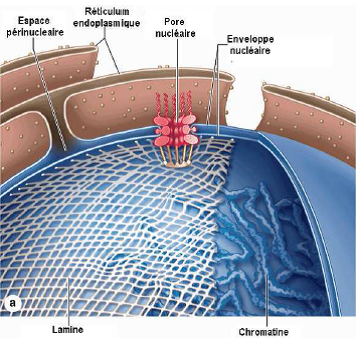
Photographie au MET d’une portion de la membrane interne de l’enveloppe nucléaire, ovocyte de xénope.

Cellule de mammifère, microscopie à super résolution
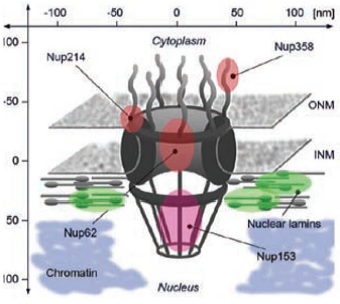
α = anticorps ; NPC = nuclear pore complex ; Nup = nucléoporine
Les lamines sont associées à l’enveloppe par des protéines transmembranaires.
Les lamines A et C sont issues du même gène.
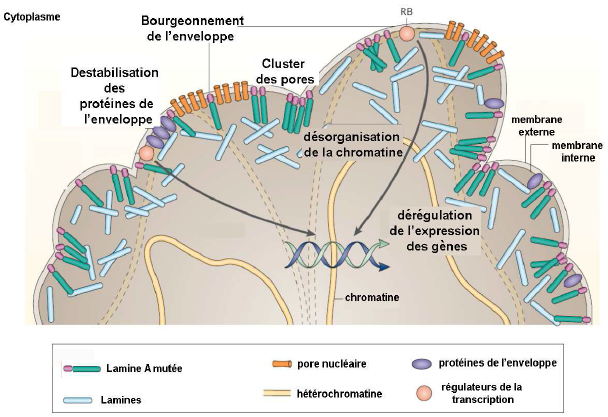
Mutation de la lamine A dans le cas du syndrome Hutchinson-Gilford Progeria (HGPS, vieillissement précoce) : mécanismes possibles.
1.6. Chromatine
Complexe formé d’ADN, d’histones et de protéines non-histones, présent dans le noyau d’une cellule eucaryote.
Ce qui forme le chromosome.
Remarque : ADN aussi dans les mitochondries.
Chez l’Homme : génome 2,85 billions de nucléotides, 46 chromosomes => 2,5 mètres => Taille du noyau ? seulement quelques micromètres
Cellule de mammifères, MET
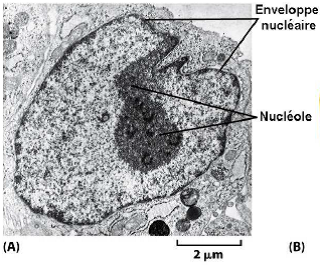
Schéma représentatif
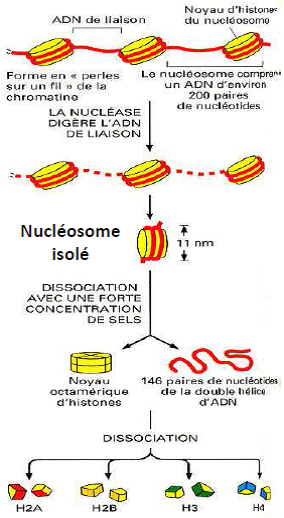
Technique d’étalement moléculaire
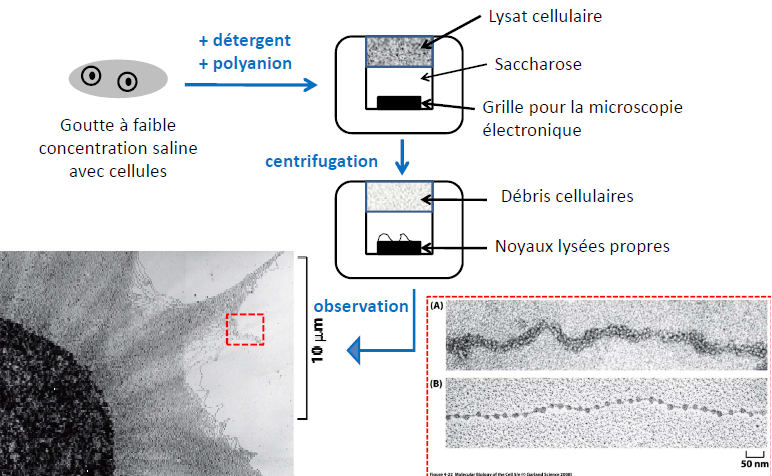
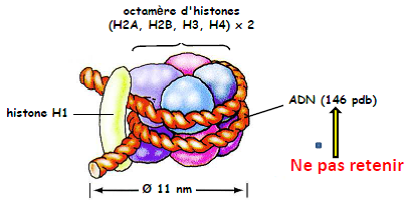
Le nucléofilament : constitué de nucléosomes successifs compactés.
Les mêmes molécules mais avec des niveaux de compaction différents.
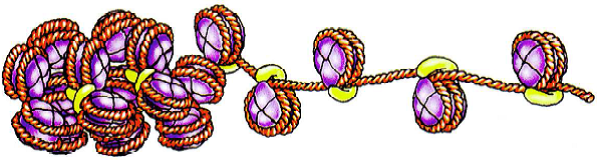
Schéma du nucléosome isolé
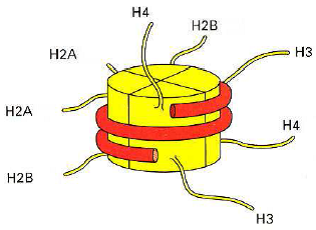
Le nucléosome = structure de base de la chromatine.
- Interactions électrostatiques => Maintien de l’ADN autour de l’octamère d’histones.
- Extrémités N terminales des histones : => repliement de l’ADN => remodelage de l’ADN (transcription, réplication, réparation).
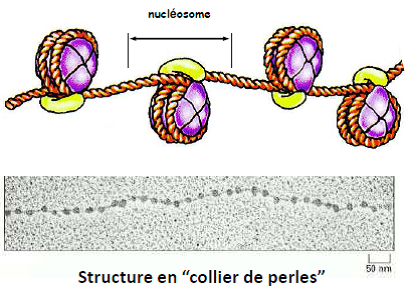
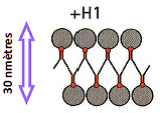
Structure en solénoïde (~ 30 nm)

Structure en « collier de perles » (~ 11 nm)
Structure en solénoïde
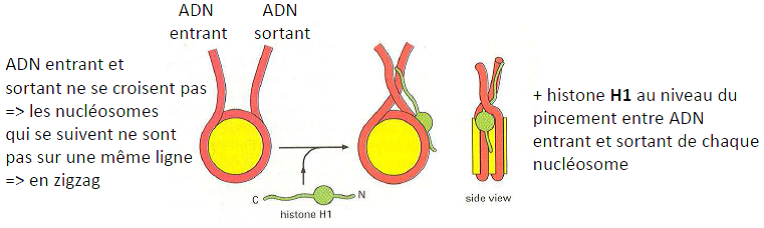
Hypothèse : H1 permettrait de déplacer les queues de H3, les rendant libérées de leur interaction avec l’ADN.
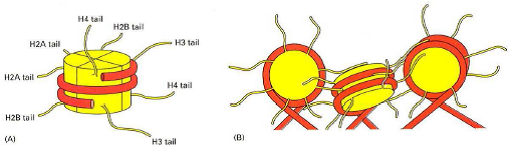
Modèle de l’arrangement en zigzag
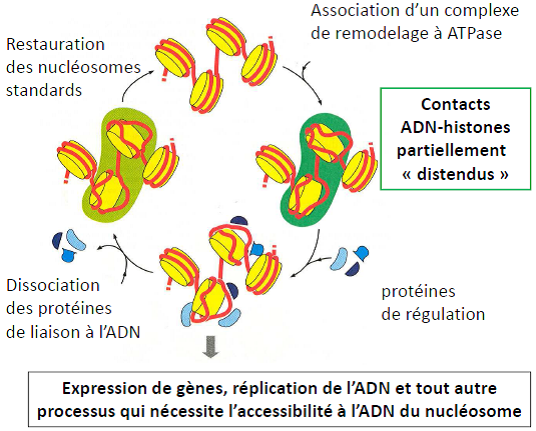
Nucléosome joue aussi un rôle essentiel dans les grandes fonctions nucléaires (transcription, réplication, réparation de l’ADN).
Modulation de l’activité du nucléosome :
- Complexes de remodelages avec ATPase.
- Les modifications post-traductionnelles (extrémités) des histones de l’octamère.
- Incorporation de variants d’histones.
Chromatine plus ou moins dense aux électrons. Cellules de mammifère, MET.
=> Niveaux de compaction variables dans un noyau.
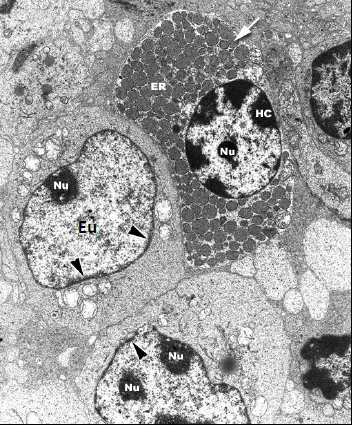
HC : Hétérochromatine
Eu : Euchromatine
Exemples de cellules plus ou moins riches en euchromatine :
Noyau riche en hétérochromatine
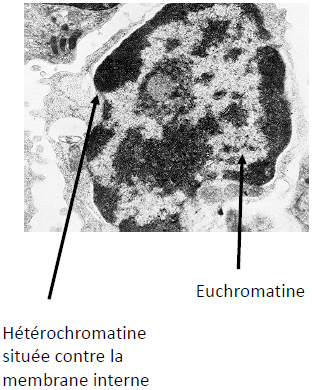
Noyau riche en euchromatine

| Hétérochromatine – Chromatine +/- condensée. – Partiellement périphérique. 2 types : – Constitutive (centromères, télomères, chromosome X) : pas transcrite. – Facultative : pas transcrite mais avec capacité de se transformer en euchromatine. | Euchromatine – Chromatine +/- décondensée. – Dispersée dans le nucléoplasme. Remarque : Aucune observation au microscope ne permet d’établir le degré de condensation de l’ADN. |
La localisation de l’hétérochromatine et de l’euchromatine peut avoir une fonction :
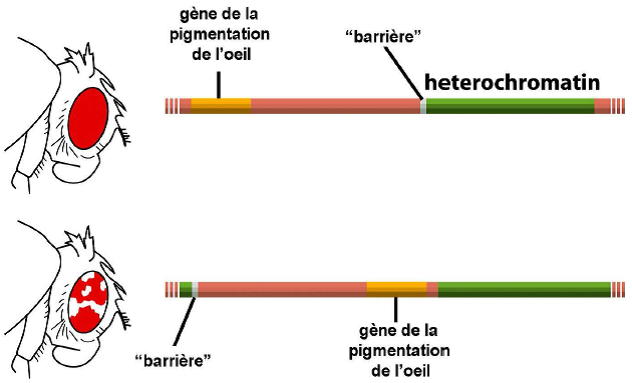
Territoires chromosomiques
FISH : hybridation in situ fluorescente (observation de séquences nucléotidiques).
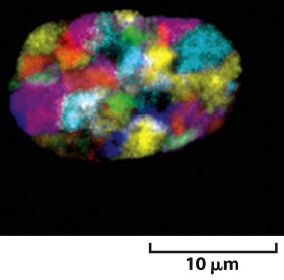
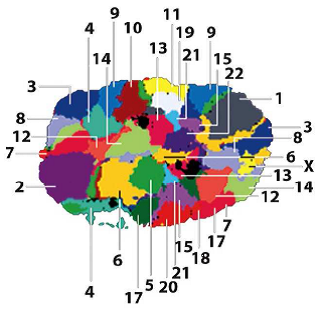
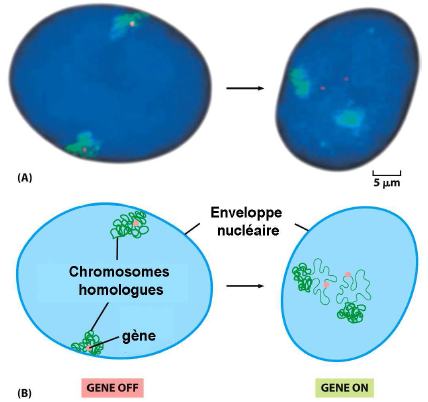
Remarque : La chromatine n’est pas « figée » dans le noyau.
=> Déplacement vers des territoires nucléaires pour l’expression des gènes.
Noyau d’une cellule humaine
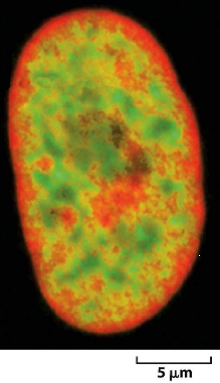
En rouge : chromatine dépourvue de « gènes ».
Où est préférentiellement localisée l’hétérochromatine ?
=> en périphérie (en rouge)
Chromosome interphasique / chromosome métaphasique
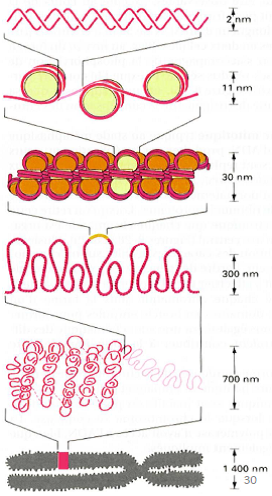
Chromosome interphasique (suivre l’ordre des 4 premières lignes de l’image ci-dessus):
- Courte région de la double hélice d’ADN
- Chromatine en « collier de perles » (succession de nucléosomes).
- Fibre chromatinienne de 30 nm constituée de nucléosomes empilés en spirale.
- Partie étalée d’un chromosome interphasique.
Chromosome métaphasique (suivre l’ordre des 2 dernières lignes de l’image ci-dessus):
- Partie condensée d’un chromosome métaphasique.
- Chromosome métaphasique entier.
Chromosomes interphasiques 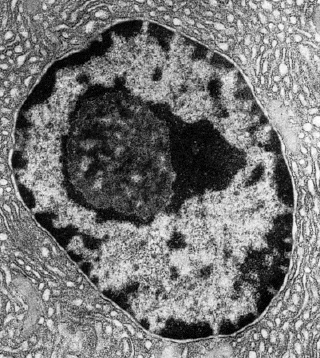 | Chromosomes mitotiques (vue caryotype)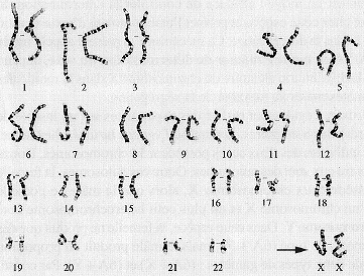 |
ME à Balayage 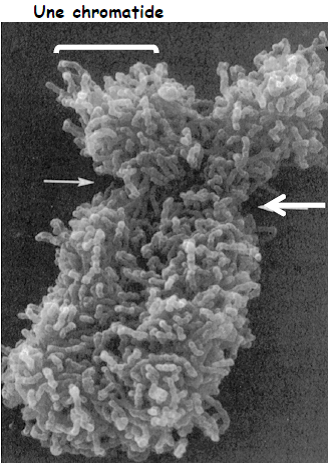 | Le chromosome mitotique est dupliqué et condensé. Télomère : extrémité d’un chromosome, avec une séquence caractéristique et répliquée spécifiquement. Centromère (flèche blanche) : Région « étranglée » d’un chromosome mitotique qui maintient les deux chromatides sœurs ensemble. Région où le kinétochore se forme. Contenu en ADN d’un chromosome eucaryote. 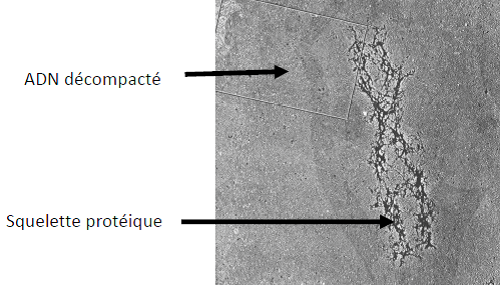 |
1.7. Nucléole
| Structure dans le noyau où l’ARNr est transcrit et les sous-unités des ribosomes sont assemblées. Remarque : Aucune membrane entoure le nucléole ! | 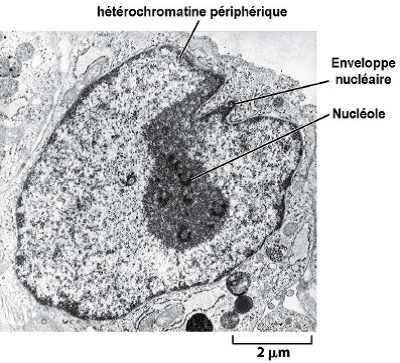 |
| Le nucléole est organisé autour des régions (boucles) chromosomiques appelées Organisateurs Nucléolaires (O.N.) qui comportent les gènes des ARNr 47S (ces régions correspondent à l’ADN des constrictions secondaires que l’on peut voir sur certains chromosomes mitotiques). | 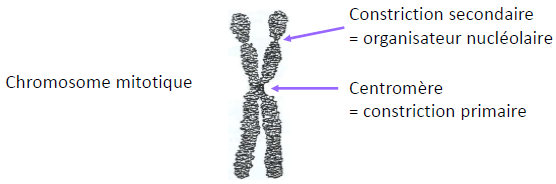 |
| Chez l’Homme : les gènes d’ADNr sont situés sur les chromosomes 13, 14, 15, 21 et 22. Il existe donc au total 10 boucles = 10 organisateurs nucléolaires dans un nucléole de cellule humaine. |  |
2. Fonctions
Expression des gènes : la transcription(voir UE 1 : fiche biochimie)
Chromosome 22 humain, sous sa conformation mitotique, composé de 2 molécules d’ADN, chacune avec 48.106 paires de nucléotides.
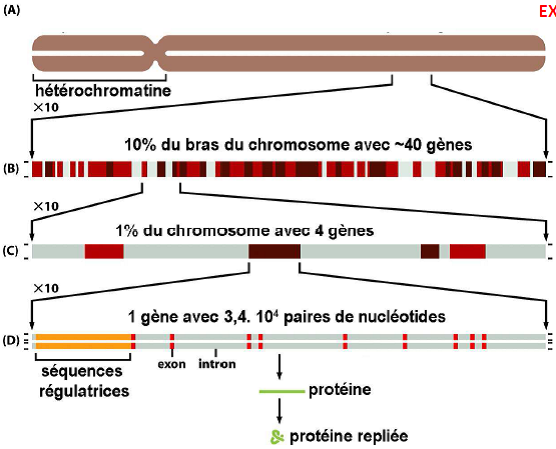 |  |
| Quel est le sens d’élongation de la transcription ? => de gauche à droite, car l’ARNm est de plus en plus long | 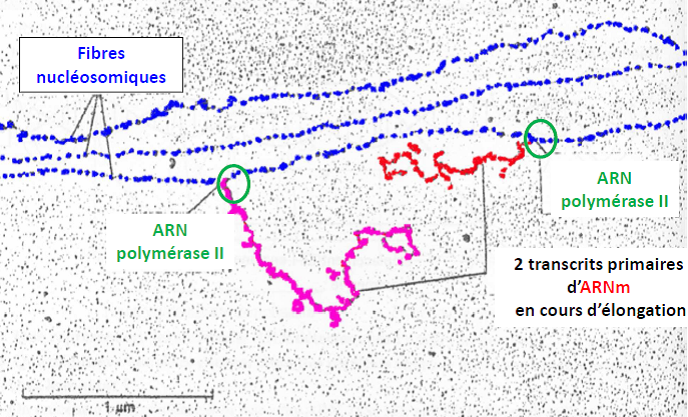 |
| Si plusieurs ARN polymérases « travaillent » à la chaîne sur le même gène, on obtient « des sapins de Noël » : | Etalement moléculaire du nucléole (synthèse des ARN ribosomiques)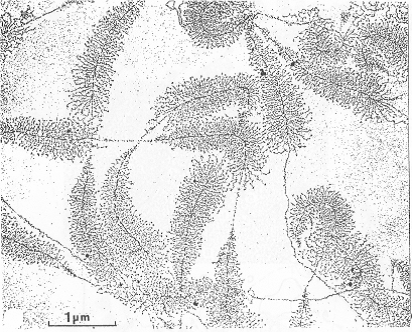 |
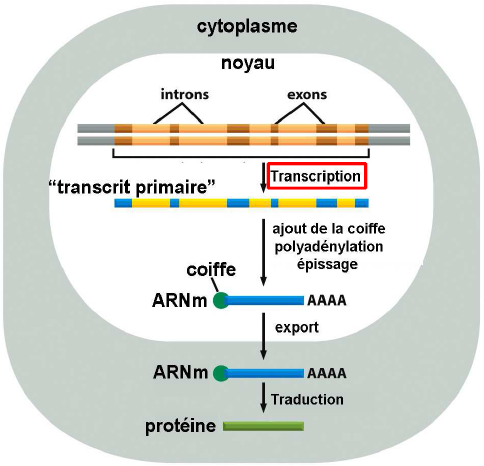
| Transcription & maturation des ARNm | 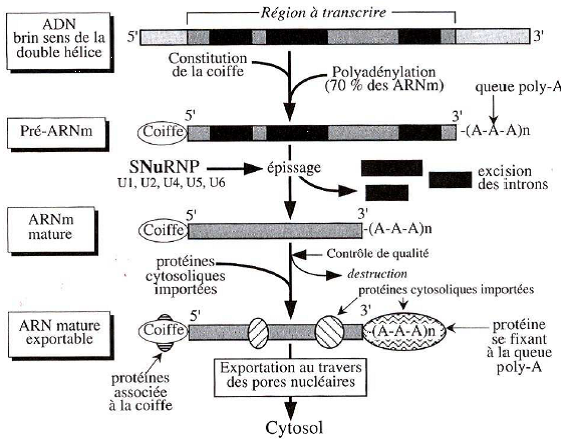 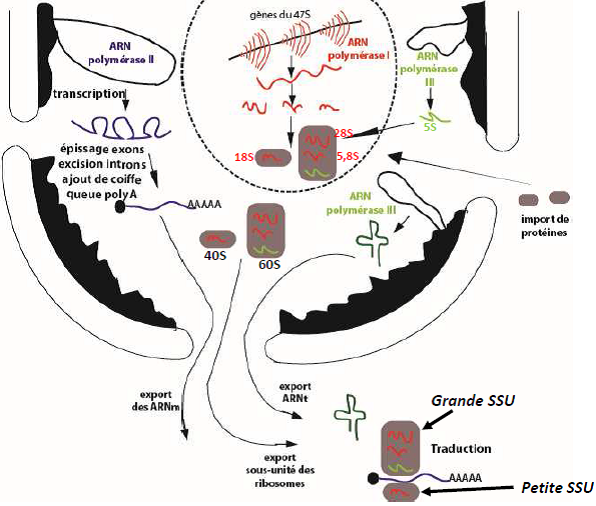 |
| Centre fibrillaire (CF) = espaceurs intergéniques non transcrits. Composant fibrillaire dense (CFD) : site de maturation des ARNr. => La transcription se déroule à la frontière entre CF et CFD. Composant granulaire : stockage des particules pré-ribosomiques avant leur exportation vers le cytosol, et de diverses protéines ribosomales. | 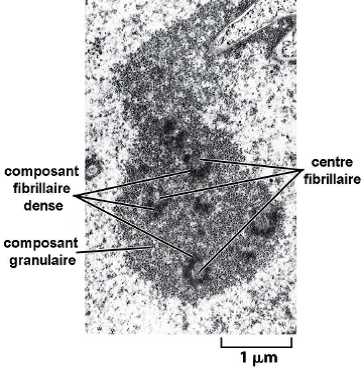 |
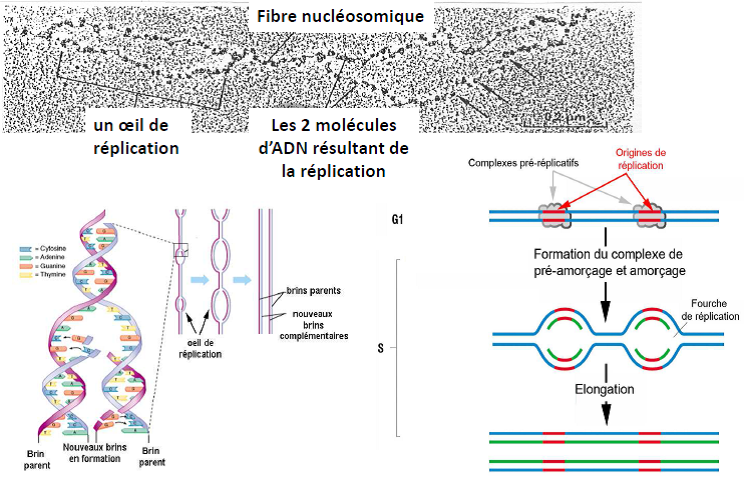
Réplication
Etalement moléculaire à partir d’une cellule en phase S
Transports nucléo-cytoplasmiques
| Cellules de mammifères, injection d’une molécule de 19kDa (vert) ou d’une protéine (rouge) | 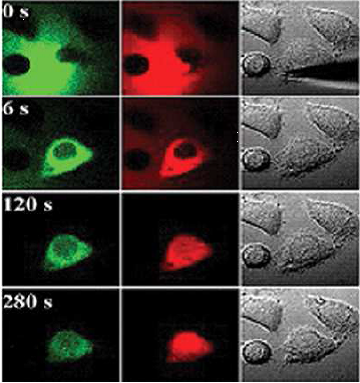 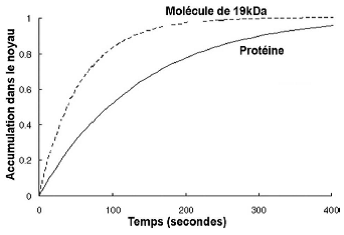 |
| Quelle est la taille : – du pore ? – d’un ARNt ? – d’un ribosome ? – d’un ARNm ? – d’une ARN polymérase ? | 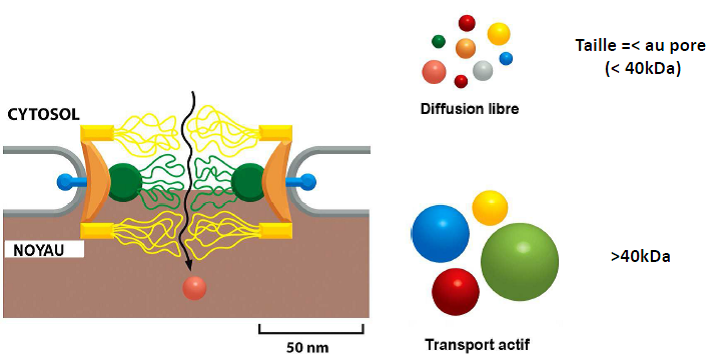 |
Cellule transfectée avec un plasmide codant une protéine « cytosolique » ; immunofluorescence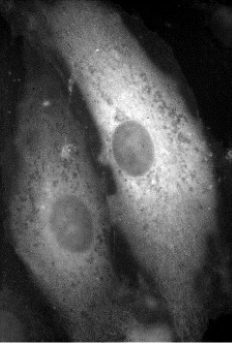 | Cellule transfectée avec un plasmide codant une protéine « cytosolique + séquence signal NLS » ; immunofluorescence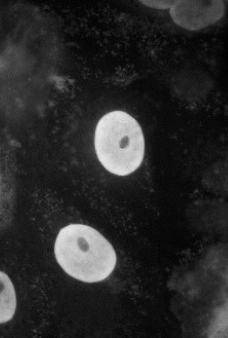 |
La séquence NLS (Nuclear Localization Signal) permet de localiser la protéine dans le noyau. Il existe un signal d’exportation du noyau = NES.
| Importation d’une protéine 1- Le cargo interagit avec le récepteur d’importation. 2- Interaction avec des acides aminés (FG) sur les filaments et les nucléoporines le long du pore nucléaire. 3- Passage dans le noyau. 4- Interaction du récepteur avec Ran-GTP & libération du cargo. 5- Ran-GTP & Récepteur d’importation interagissent avec les nucléoporines. 6- Passage dans le cytosol. 7- Hydrolyse du GTP en GDP+Pi et largage du récepteur d’import. | 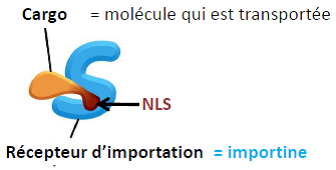 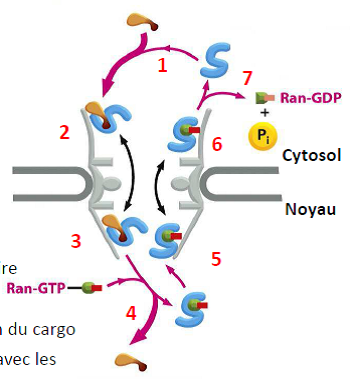 |
| Ran : petite protéine G pouvant lier le GTP (guanine tri-phosphate ou le GDP (di-phosphate) => modifie l’affinité pour le récepteur d’importation. Quelle est la fonction de Ran ? Quelle est la conséquence de la localisation de Ran-GEF dans le noyau ? | 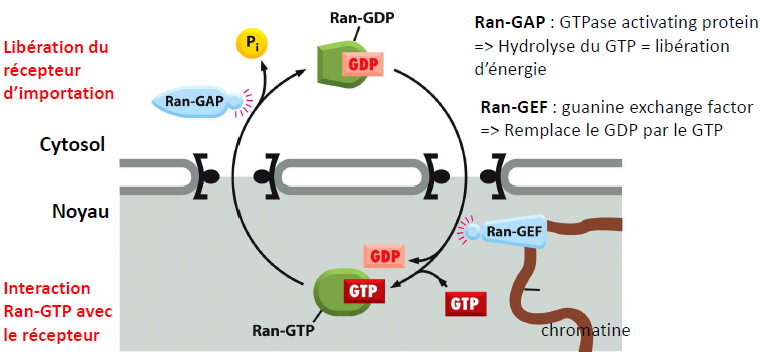 |
| Exportation d’une protéine 1- Le cargo portant un NES interagit avec Ran-GTP et le récepteur d’exportation. 2- Interaction avec les nucléoporines du pore nucléaire. 3- Sortie du noyau. 4- Hydrolyse du GTP et libération du cargo dans le cytosol. 5- Le récepteur d’exportation interagit avec les nucléoporines. 6- Le récepteur d’exportation retourne dans le noyau. Comment le Ran-GDP entre dans le noyau ? | 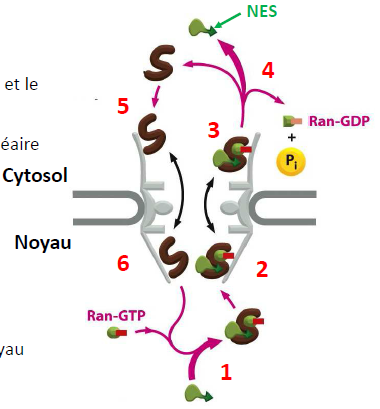 |
| Transport de Ran-GDP du cytoplasme vers le noyau NTF2 : famille de récepteur du transport nucléaire différente des importines. | 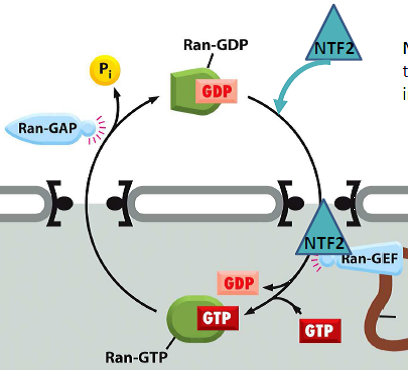 |
| Exportation de l’ARNm : ARNt et ARNr/Ribosomes utilisent le Ran-GTP. | 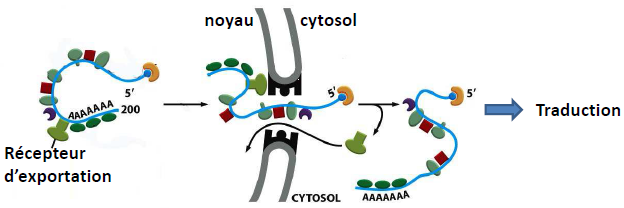 |